Mutagenese: wie entsteht eine Mutation?
Mutationen unterteilt man nach Art ihrer Entstehung in spontane und induzierte Mutationen. Spontane Mutationen geschehen auch unter "normalen" Verhältnissen mit bestimmter Wahrscheinlichkeit. Die Spontanrate ist allerdings individuell verschieden. Der Grund für diese Variation liegt in der unterschiedlichen Aktivität der DNA-Reparatursysteme (siehe weiter unten). Die Mutationsrate wird durch induzierte Mutationen weiter erhöht. Physikalische, chemische oder auch virale Mutagene induzieren Mutationen.
Die Grenze zwischen spontanen und induzierten Mutationen ist aber nicht immer einfach zu ziehen: Viele Agentien müssten nämlich eigentlich in beiden Klassen aufgeführt werden, weil eine gewisse Dosis eines Mutagens auch unter normalen Bedingungen existiert. Ultraviolette Strahlung, zum Beispiel, ist auch im Spektrum des Sonnenlichts enthalten und ist so auch an der Entstehung der "spontanen" Mutationen beteiligt.
Einen Spezialfall stellen genetisch programmierte Mutationen dar. Beispiel: Induzierte Mutationen in Genen, die für variable Anteile von Antikörpern kodieren. Das geschieht in B-Zellen, die ihre Antikörper-Gene rearrangiert haben und mit einem passenden Antigen in Kontakt kommen. Mechanismus: Aktivierung einer Cytosin-Deaminase. Biologischer Sinn: Bildung von Antikörpern mit höherer Affinität für das Antigen.
Induzierte Mutationen
1. Physikalische Mutagene
Ionisierende Strahlen
- Ionisierende Strahlen sind energiereiche Strahlen (Röntgenstrahlen, Alpha- Beta- oder Gamma- Strahlen radioaktiver Substanzen), die einen Teil der von ihnen getroffenen Moleküle verändern. Röntgenstrahlen z.B. rufen bevorzugt Chromosomenbrüche hervor.
- UV-Bestrahlung führt zur Bildung von Dimeren aus zwei benachbarten Pyrimidin-Resten, wie weiter unten genauer beschrieben wird.
2. Chemische Mutagene
ENU und EMS
- Ethylnitroisoharnstoff (ENU) und Ethylmethansulfonat (EMS) werden experimentell zur Mutagenese eingesetzt. ENU und EMS lösen Punktmutationen aus.
Colchizin
- Polyploidie kann durch Zugabe von Colchizin induziert werden. Colchizin hemmt nämlich den Spindelapparat und damit die Trennung der Chromatiden bei der Zellteilung, indem es an Tubulin bindet und die Neubildung von Mikrotubuli verhindert.
5-Brom-Uracil
- 5-Bromuracil ist eine dem Thymin ähnliche Base - sie trägt ein Bromatom anstelle der Methylgruppe. Bei der Replikation wird es aber oft mit Cytosin verwechselt, so dass Guanin anstelle von Adenin in die DNA eingebaut wird. Dies ergibt schliesslich eine Basensubstitution.
- Salpetrige Säure (HNO2) ist Katalysator bei diversen Umwandlungsreaktionen: So entstehen durch Abspaltung von NH3 (Desaminierung) und anschliessender Isomerisierung aus Cytosin Uracil, aus Adenin Hypoxanthin und aus Guanin Xanthin. (Formeln zeigen)XSalpetrige Säure
Salpetrige Säure (HNO2) ist Katalysator bei diversen Umwandlungsreaktionen: So entstehen durch Abspaltung von NH3 (Desaminierung) und anschliessender Isomerisierung aus Cytosin Uracil, aus Adenin Hypoxanthin und aus Guanin Xanthin, wie untenstehende Formeln zeigen: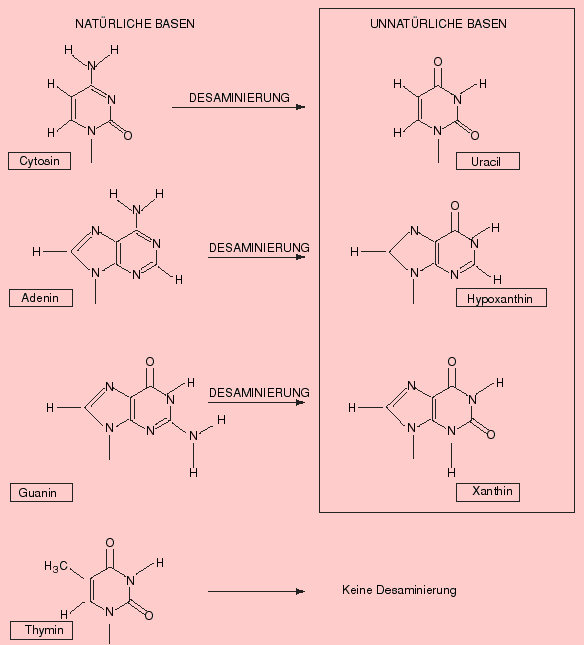 Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990, ISBN 3-527-27983-0
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990, ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
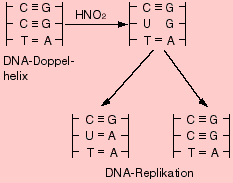
Eine der beiden Cytosin-Basen im DNA-Abschnitt wird unter Einfluss von HNO2 desaminiert. Welche Folgen hat dies auf diesen DNA-Abschnitt und welche Basensequenz haben die bei der nachfolgenden Replikation entstehenden DNA-Helices?
(Tipp: Uracil paart normalerweise mit Adenin)
Lösung zur Aufgabe
X
Virale Mutagene
Auch Viren
X
VirenViren sind infektiöse Partikel, die sich in eukaryotischen Zellen vermehren. Viele von ihnen sind als Krankheitserreger bei Menschen, Tieren und Pflanzen medizinisch und ökonomisch von Bedeutung. Aehnlich wie Bakteriophagen in prokaryotischen Zellen sind Viren in eukaryotischen Zellen bedeutsame Modellsysteme für molekularbiologische Studien; einige von ihnen dienen zusätzlich als Vektoren für Gentransferexperimente in eukaryotischen Systemen. Wie die Bakteriophagen sind auch Viren zur Vermehrung auf eine lebende Wirtszelle angewiesen.
Strukturen
In Form, Grösse, Informationsgehalt des Genoms, Wirtspezifität etc. herrscht eine ungeheure Vielfalt. Wir werden deshalb im folgenden nur einige grundlegende Eigenschaften besprechen und dabei das eine oder andere Virus als Beispiel heranziehen (siehe Tabelle).

Viren bestehen in der Regel aus folgenden Komponenten:
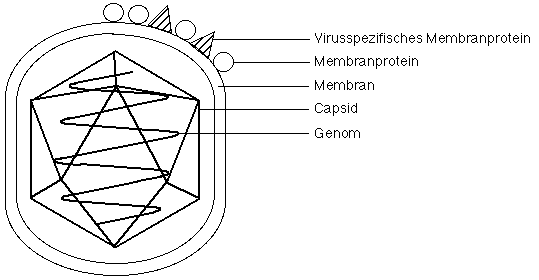
Genom
Virale Genome können aus doppelsträngiger (ds) DNA, einzelsträngiger (ss) DNA, dsRNA oder RNA bestehen. Ihre Grössen variieren zwischen 10 hoch 3 (Parvovirus) bis mehr als 10 hoch 6 Nukleotide (Poxvirus). Genome können als lineare, zirkuläre oder fragmentierte Nukleinsäure-Moleküle vorliegen. Bei einzelsträngigen Nukleinsäuren kann der Strang mit positiver Polarität (gleiche Sequenz wie mRNA) oder der Strang mit negativer Polarität als Genom dienen; für eine bestimmte Virus-Art ist es aber stets derselbe Strang. Virale Genome kodieren für Strukturproteine des Virus und für Enzyme, die für die Genom-Replikation benötigt werden.
Capsid
Eine Proteinhülle, das Capsid, umgibt das virale Genom. Das Capsid ist aus Struktureinheiten, den Capsomeren, zusammengesetzt, die in der Regel aus einer grossen Zahl eines einzigen oder ein paar wenigen Typen von Proteinen bestehen. Oft sind die Capsomere, wie in der Abbildung dargestellt, als Icosaeder angeordnet (20 gleichseitige Dreiecke). Viele Viren, unter ihnen das Poliovirus (siehe Tabelle), bestehen nur aus Genom und Capsid.
Membran
Sie ist, wie vorher angedeutet, nicht bei allen Viren vorhanden. Die Membran stammt von der Wirtszelle und wird vom Virus beim Austritt aus der Zelle mitgenommen (siehe unten). Sie enthält in der Regel vom Wirtsgenom kodierte und eventuell zusätzlich virus-kodierte Proteine. Beispiele von Viren mit einer Membran sind (in der Tabelle) Influenzavirus und die Retroviren.
Einteilung der Viren
Viren werden aufgrund ihrer Morphologie und der Grösse und Art ihres Genoms klassifiziert.
Vermehrung
Eintritt in die Zelle:
Viren adsorbieren an spezifische Oberflächen-Moleküle der Wirtszelle. Es ist somit die Oberflä-chenstruktur einer Zelle, die mitentscheidet, ob eine Infektion mit einem bestimmten Virus möglich ist. Die Aufnahme eines adsorbierten Virus in die Zelle geschieht dann via Endocytose. Der Inhalt des infizierenden Virus wird darauf in der Zelle entlassen.
Expression und Replikation des viralen Genoms:
Die meisten Viren, die dsDNA als Genom besitzen, replizieren und transkribieren die DNA im Zellkern der Wirtszelle mit Hilfe wirts-spezifischer Enzyme. Eine Ausnahme bildet das Poxvirus, das sich im Cytoplasma vermehrt und alle Enzyme für Replikation und Transkription mitbringt.
Viren mit RNA-Genomen von positiver Polarität (z.B. Polioviren) können direkt mit der Proteinsynthese im Cytoplasma beginnen, da ihr Genom als mRNA dienen kann. Sie replizieren ihr RNA-Genom mit Hilfe einer virus-spezifischen Replicase, welche zuerst eine RNA-Kopie mit negativer Polarität synthetisiert und diese anschliessend in RNA-Genome mit positiver Polarität kopiert. Alle diese Vorgänge finden im Cytoplasma statt.
Viren mit RNA-Genomen von negativer Polarität (z.B. Influenzavirus) oder dsRNA (z.B. Reovirus) benötigen eine RNA-abhängige RNA-Polymerase für Transkription und Replikation und müssen diese, da die Wirtszellen kein solches Enzym besitzen, im Viruspartikel mit in die Zelle bringen.
Retroviren haben einen besonders interessanten Mechanismus der Replikation und Transkription entwickelt. Sie überschreiben ihr RNA-Genom mit positiver Polarität mittels einer virus-kodierten Reversetranskriptase in dsDNA. Diese dsDNA-Kopie des viralen Genoms wird in die Wirtszell-DNA integriert und dort wie zelleigene Gene von wirts-spezifischen Enzymen transkribiert.
Synthese der viralen Proteine
Allen Viren ist gemeinsam, dass sie zur Uebersetzung ihrer mRNA vollständig auf den Protein-syntheseapparat der Wirtszelle angewiesen sind. Diese Synthese findet daher immer im Cytoplasma statt. Der darauffolgende Zusammenbau der Komponenten zum intakten Nachkommenvirus findet, je nach Virusart, im Cytoplasma oder im Zellkern statt.
Freisetzung der Viren
Nachkommenviren können am Ort des Zusammenbaus riesige intrazelluläre Aggregate bilden, welche nach dem Absterben und Lysieren der Wirtszelle freigesetzt werden. Gewisse Viren (z.B. Influenzavirus) synthetisieren virus-spezifische Membranproteine, welche in die Plasmamembran der Wirtszelle integrieren und später das Capsid binden. Das Nachkommenvirus tritt dann durch die Membran aus, wobei es ein Stück dieser Membran mitnimmt:

Spontane Mutationen
Desaminierungen und Depurinierungen
Dass salpetrige Säure die Struktur von Basen ändern kann, wurde schon oben gezeigt. Diese Desaminierung kann aber auch spontan ablaufen, genauso wie die hydrolytische Spaltung der Zucker-Base-Bindung von Purinen (Depurinierung) (Formeln zeigen)
X
Desaminierungen und DepurinierungenDass salpetrige Säure die Struktur von Basen ändern kann, wurde schon oben gezeigt. Diese Desaminierung kann aber auch spontan ablaufen, genauso wie die hydrolytische Spaltung der Zucker-Base-Bindung von Purinen (Depurinierung):
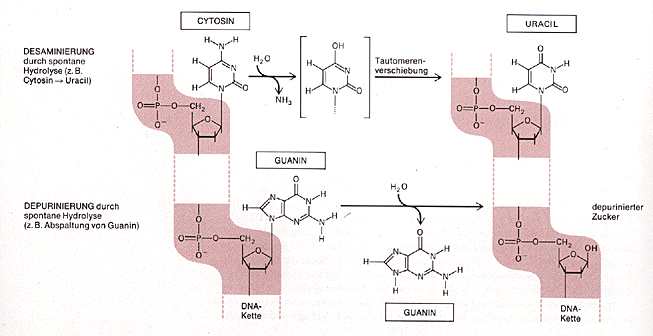 Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990, ISBN 3-527-27983-0
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990, ISBN 3-527-27983-0und 3. Auflage 1995, ISBN 3-527-30055-4
Pyrimidin-Dimere
Benachbarte Pyrimidin-Basen (T-T, T-C, C-C) werden durch ultraviolette Strahlung zu kovalent gebundenen Pyrimidin-Dimeren verbunden. Beispiel: Thymin-Dimer
X
Benachbarte Pyrimidin-Basen (T-T, T-C, C-C) werden durch ultraviolette Strahlung zu kovalent gebundenen Pyrimidin-Dimeren verbunden, wie hier am Beispiel des Thymin-Dimers zu sehen ist: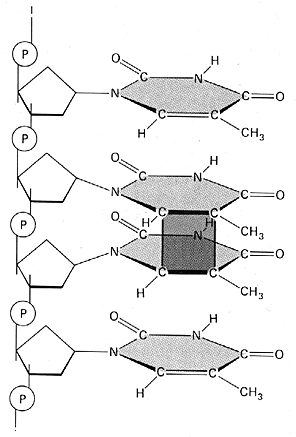 Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH,
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH,2. Auflage 1990, ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Endoreplikation, Zellkernfusion und Endomitose
Polyploide Zellen entstehen entweder durch doppelte DNA-Synthese in der S-Phase des Zellzyklus (Endoreplikation), durch Fusion von Zellkernen (Zellkernfusion) oder durch DNA-Synthese unter Ausbleiben der Zell- und Kernteilung (Endomitose).
Non-disjunction
Wenn sich zwei homologe Chromosomen bei der Meiose nicht trennen, erhält die eine Keimzelle beide Chromosomen, die andere keines. Diese Erscheinungsform nennt man meiotische Non-disjunction. Nichttrennung von homologen Chromatiden in der Mitose (mitotische Non-disjunction) hat zur Folge, dass ein Chromosom zuviel oder zuwenig in eine somatische Zelle gelangt. Beide Kategorien von Non-disjunction führen schliesslich zur Aneuplodie: Die Zahl der Chromosomen einer Zelle weicht von 23 Chromosomenpaaren ab.
Tautomerien
Die in der DNA vorkommenden Basen tragen unter anderem Amino- und Ketogruppen. Sie können aber in seltenen Fällen intramolekulare Umlagerungsreaktionen erfahren und in der Imino- oder Enolform vorliegen. In diesem Fall ändern sich die Paarungseigenschaften: So paart z.B. Cytosin nicht mehr mit Guanin, sondern mit Adenin! (Formel zeigen)
X
TautomerienDie in der DNA vorkommenden Basen tragen unter anderem Amino- und Ketogruppen. Sie können aber in seltenen Fällen intramolekulare Umlagerungsreaktionen erfahren und in der Imino- oder Enolform vorliegen. In diesem Fall ändern sich die Paarungseigenschaften: So paart z.B. Cytosin nicht mehr mit Guanin, sondern mit Adenin!
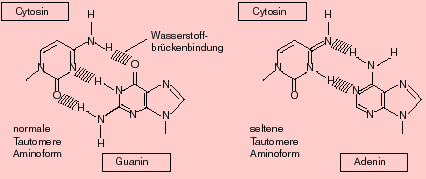 Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990,
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990,ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Entsprechendes gilt für alle andern Basen, wie folgende Aufstellung zeigt:
| Normale Form | Tautomere Form |
| A (Amino) paart mit T | A (Imino) paart mit C |
| C (Amino) paart mit G | C (Imino) paart mit A |
| G (Keto) paart mit C | G (Enol) paart mit T |
| T (Keto) paart mit A | T (Enol) paart mit G |
Zu den Übungsaufgaben