Reparaturmechanismen
Zellen verlieren durch spontane Depurinierung pro Tag bis zu 5000 Purinbasen aus der DNA. Die spontanen Desaminierungen von Cytosin zu Uracil werden auf etwa 100 geschätzt. Das Verhältnis zwischen den bekannten DNA-Basen und ihren tautomeren Formen beträgt etwa 10'000 zu 1. Trotzdem wird bei der Replikation nur etwa eine Base auf 10 hoch 9 Basen falsch eingebaut. Wie ist dieser Widerspruch zu erklären? Warum macht ein durchschnittlich grosses Gen nur alle 10 hoch 6 Zellteilungen eine Mutation durch?
Obige Zahlen fordern die Existenz von Reparatursystemen, welche die DNA nach Fehlern absuchen und diese korrigieren. Solche Reparatursysteme gibt es tatsächlich. Als Mutationen erscheinen also nur die Genomänderungen, welche von den Reparaturkomplexen nicht erkannt wurden.
Zellen verfügen über eine Reihe von Enzymkomplexen, welche verschiedene Typen von DNA-Schäden erkennen und reparieren können. Vereinfacht bestehen DNA-Reparaturvorgänge aus drei Schritten:
- Ein Fehler auf einem DNA-Abschnitt wird von einem Enzymkomplex erkannt. Endonukleasen hydrolysieren darauf die Phosphodiesterbindungen im fehlerhaften Abschnitt und entfernen so die veränderte Nukleotidsequenz.
- Der resultierende DNA-Einzelstrang wird durch eine DNA-Polymerase wieder mit komplementären Basen zum Doppelstrang ergänzt.
- Eine DNA-Ligase verbindet das zuletzt eingebaute Nukleotid mit seinem Nachbarn und schliesst damit die Lücke.
Depurinierte Stellen
- Eine Endonuklease spaltet die DNA-Kette gerade vor der defekten Stelle und entfernt den Zucker und das Phosphat.
- Eine DNA-Polymerase setzt das passende Nukleotid ein.
- Eine DNA-Ligase schliesst die Lücke
Enzyme erkennen Veränderungen in der DNA-Doppelhelix
Fehler in der DNA-Doppelhelix, welche die Basenpaarung verändern, werden von Glycosylasen erkannt und die entsprechenden Basen entfernt. Glycosylasen sind eine vielfältige Gruppe von Enzymen: es gibt Glycosylasen, die spezifisch desaminierte Adenine, desaminierte Cytosine, verschiedene alkylierte Basen und Basen mit geöffneten Ringen entfernen können. Nach dem Entfernen der defekten Base wird der Schaden wie oben beschieben repariert.
Desaminierungen können erkannt werden, weil jede durch eine Desaminierung entstandene Base nicht mehr korrekte Basenpaarung eingehen kann: so wird aus Cytosin Uracil, aus Adenin Hypoxanthin und aus Guanin Xanthin. Hier liegt ein Evolutionsvorteil, den die Verwendung von Thymin anstelle von Uracil im Einbau in die DNA hat: in einer uracil-haltigen DNA könnten normale Uracile nicht von solchen unterschieden werden, die durch Desaminierung von Cytosin entstehen.
Es gibt auch Multienzymkomplexe, die grössere Veränderungen in der DNA erkennen und reparieren können. Solche Schäden entstehen unter anderem bei der Anlagerung von Kohlenwasserstoffen (wie z.B. dem Karzinogen Benzpyren) an Basen oder Bestrahlung von DNA mit ultravioletter Strahlung, welche zu kovalenter Verknüpfung von benachbarten Pyrimidinen führt. Solche DNA-Stränge werden vor und nach der schadhaften Stelle von Endonukleasen gespalten, ein Stück DNA von 20-30 Nukleotiden mit der schadhaften Stelle entfernt. Die entstandene Lücke wird darauf wie oben beschrieben repariert.
Und wenn beide DNA-Stränge defekt sind ?
Sind gleichzeitig beide Kopien am gleichen Basenpaar beschädigt, kann keiner der beiden Stränge mehr als Matrize für die Reparatur gebraucht werden. Aber auch für diesen seltenen Fall gibt es mittels genetischer Rekombinationsvorgänge manchmal eine Rettung, nämlich dann, wenn die defekte Basensequenz noch in einem andern DNA-Abschnitt unversehrt vorhanden ist.
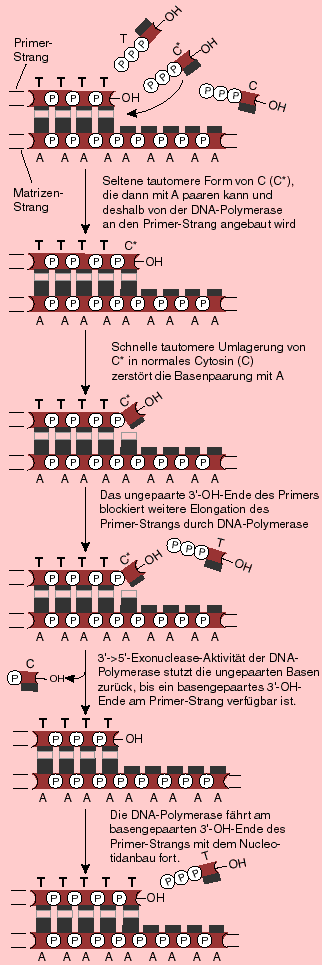
Der "Korrekturlese"-Mechanismus der DNA-Polymerasen
Die DNA-Polymerase baut bei der Replikation Nukleotide in 5'-> 3' - Richtung in den wachsenden DNA-Strang ein. Dazu braucht sie ein korrektes Basenpaar und ein freies 3' -OH-Ende, an welches das nächste Nukleotid angehängt werden kann. Liegen nun zwei nicht-komplementäre Basen vor (wie es beim Einbau von tautomeren Formen der Basen geschehen kann), stoppt die Polymerase und entfernt solange Nukleotide in 3'-> 5' - Richtung, bis wieder ein basengepaartes Ende zur Verfügung steht.
Die DNA-Polymerase ist also ein selbst-korrigierendes Enzym, indem es die von ihr falsch eingebauten Basen erkennt und wieder entfernt.
Abbildung
X
Der "Korrekturlese"-Mechanismus der DNA-PolymerasenDie nebenstehende Abblildung zeigt, wie die seltene tautomere Form von Cytosin von der Exonuclease-Domäne der DNA-Polymerase erkannt wird und durch die richtige Base Adenin ersetzt.
 Bildquelle: Alberts et al,: "Molekularbiologie der Zelle",
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle",VCH, 2. Auflage 1990, ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Zu DNA-Polymerase (im Kapitel "Mechanismus der Replikation")
Zu den Übungsaufgaben
Die Reparaturmechanismen für eine Zelle sind sehr wichtig. Patienten mit der Erbkrankheit Xeroderma pigmentosum haben Defekte in gewissen Reparaturenzymen und leiden unter schweren Hautschäden, darunter auch Hautkrebs.